![]() To English
To English
![]() To Arabic-English
To Arabic-English
![]() To Arabic
To Arabic
Vision
Ranges Linked to Quran and Hadith
Hussain Omari
Physics Dept./ Mutah University/ Jordan
rashed@mutah.edu.jo
الملخص:
قال رسول
اللّه صلى
اللّه عليه
وسلّم: (إذا
سمعتم صياح
الديكة
فاسألوا الله
من فضله ، فإنها
رأت ملكا ،
وإذا سمعتم
نهيق الحمار
فتعوذوا
بالله من
الشيطان ،
فإنه رأى
شيطانا) . يبين
المقال أنّ نطاق
الرؤية (Vision Range)
يختلف من كائن
إلى آخر كما
يرشد إليه هذا
الحديث
الشريف : الديكة
ترى بعض الملك
، والحمار يرى
بعض الشياطين
، وفي
الحالتين لا يرى
الإنسان
شيئاً من هذا. وإِنَّ
الشَّيْطَان
يَرَانا هُوَ
وَقَبِيلُهُ
مِنْ حَيْثُ
لا نراهم : (إِنَّهُ
يَرَاكُمْ
هُوَ
وَقَبِيلُهُ
مِنْ حَيْثُ
لَا
تَرَوْنَهُمْ) . وثبت أنَّ
أسيدَ بنَ
حضيرٍ ، بينما
هو ، ليلةً ،
يقرأُ في
مربدِه إذ إذ
جالت فرسُه
ثلاث مرّات . ...
قال فانصرفتُ
. وكان يحيى
قريبًا منها .
خشيتُ أن
تطأَه .
فرأيتُ مثلَ
الظُلَّةِ .
فيها أمثالُ
السُّرُجِ .
عرجتْ في
الجوِّ حتى ما
أراها . فقال
رسولُ اللهِ
صلَّى اللهُ عليهِ
وسلَّمَ " تلك
الملائكةُ
كانت تستمعُ لك
. ولو قرأتَ لأصبحتْ
يراها الناسُ
. ما تستتِرُ
منهم ؛ ممّا
يشير إلى
أنّها أصلاً
مستترة عنّا
ولا يمكننا
رؤيتها.
(Narrated Abu Huraira: The Prophet said, "When you hear the
crowing of cocks, ask for Allah's Blessings for (their crowing indicates that)
they have seen an angel. And when you hear the braying of a donkey, seek Refuge
with Allah from Satan for (its braying indicates) that it has seen a
Satan." ). This hadith refers to
the scientific fact that Vision Ranges differ for different creatures: Cocks can
see some of the angels, and a donkey can see some demons, and in both cases the
human does not see anything from this. The devil and its tribe see us, while we
do not see them (He
(Satan) and his tribe watch you from a position where ye cannot see them).
Meanwhile a companion of
the Prophet reading Al-Kahf surah and beside him a horse hitched with two ropes, a rotating cloud starts approaching him and
make his horse alienate. When the day
light appears, he went to the Prophet peace be upon him and mentioned that to
him. Prophet said those tranquility descend for the
Qur-an. According to another version,
these angels were listening to you. Had
you read more, it will become apparent to people, and no more hidden.
المبحث
الأوّل : اختلاف
نطاق الرؤية
من كائن إلى
آخر كما بيّنت
الآيات
والأحاديث
الفرع
الأوّل:
الديكة ترى
بعض الملك،
والحمار يرى
بعض الشياطين
وفي الحالتين
لا يرى
الإنسان
شيئاً من هذا
جاء في
الحديث
الصحيح ، الذي
يرويه أبو
هريرة، قال
رسول اللّه
صلى اللّه عليه
وسلّم: (إذا
سمعتم صياح
الديكة
فاسألوا الله
من فضله ، فإنها
رأت ملكا ،
وإذا سمعتم
نهيق الحمار
فتعوذوا
بالله من
الشيطان ،
فإنه رأى
شيطانا) (الراوي:
أبو هريرة
المحدثون:
البخاري
- المصدر: صحيح
البخاري -
الصفحة أو الرقم:
3303، مسلم -
المصدر: صحيح
مسلم - الصفحة
أو الرقم: 2729،
أبو داود -
المصدر: سنن
أبي داود - الصفحة
أو الرقم: 5102،
الألباني -
المصدر: صحيح
الترمذي -
الصفحة أو
الرقم: 3459،
الألباني -
المصدر: صحيح
أبي داود -
الصفحة أو
الرقم: 5102).
يؤكّد
هذا الحديث
الشريف أنّ
الديكة ترى
بعض الملك ،
كما وأنّ والحمار
يرى بعض
الشياطين ، وفي
الحالتين لا
يرى الإنسان
شيئاً من هذا.
الفرع
الثاني:
إِنَّهُ
يَرَاكُمْ
هُوَ وَقَبِيلُهُ
مِنْ حَيْثُ
لَا
تَرَوْنَهُمْ
وجاء
في الآية
الكريمة: (يَا
بَنِي آدَمَ
لَا
يَفْتِنَنَّكُمُ
الشَّيْطَانُ
كَمَا
أَخْرَجَ
أَبَوَيْكُمْ
مِنَ الْجَنَّةِ
يَنْزِعُ
عَنْهُمَا لِبَاسَهُمَا
لِيُرِيَهُمَا
سَوْآتِهِمَا
إِنَّهُ
يَرَاكُمْ
هُوَ
وَقَبِيلُهُ
مِنْ حَيْثُ
لَا
تَرَوْنَهُمْ)
(الأعراف 27).
[27] O ye Children of Adam! let not Satan seduce you, in the
same manner as he got your parents out of the Garden, stripping them of their raiment,
to expose their shame: for he and his tribe watch
you from a position where ye cannot see them: We made the Evil Ones
friends (only) to those without Faith.
" قَالَ
بَعْض
الْعُلَمَاء :
فِي هَذَا
دَلِيل عَلَى
أَنَّ
الْجِنّ لَا
يُرَوْنَ ;
لِقَوْلِهِ "
مِنْ حَيْثُ
لَا تَرَوْنَهُمْ
" قِيلَ :
جَائِز أَنْ
يُرَوْا ; لِأَنَّ
اللَّه
تَعَالَى
إِذَا
أَرَادَ أَنْ
يُرِيَهُمْ
كَشَفَ
أَجْسَامهمْ
حَتَّى تُرَى
. قَالَ
النَّحَّاس : "
مِنْ حَيْثُ
لَا
تَرَوْنَهُمْ
" يَدُلّ عَلَى
أَنَّ
الْجِنّ لَا
يُرَوْنَ
إِلَّا فِي
وَقْت نَبِيّ
; لِيَكُونَ
ذَلِكَ
دَلَالَة
عَلَى
نُبُوَّته ;
لِأَنَّ
اللَّه جَلَّ
وَعَزَّ
خَلَقَهُمْ
خَلْقًا لَا
يُرَوْنَ فِيهِ
, وَإِنَّمَا
يُرَوْنَ
إِذَا
نُقِلُوا عَنْ
صُوَرِهِمْ .
وَذَلِكَ
مِنْ
الْمُعْجِزَات
الَّتِي لَا
تَكُون إِلَّا
فِي وَقْت
الْأَنْبِيَاء
صَلَوَات اللَّه
وَسَلَامه
عَلَيْهِمْ .
قَالَ
الْقُشَيْرِيّ
: أَجْرَى
اللَّه الْعَادَة
بِأَنَّ
بَنِي آدَم
لَا يَرَوْنَ
الشَّيَاطِين
الْيَوْم .
وَفِي
الْخَبَر (
إِنَّ الشَّيْطَان
يَجْرِي مِنْ
اِبْن آدَم
مَجْرَى
الدَّم ) .
وَقَالَ تَعَالَى
: " الَّذِي
يُوَسْوِس
فِي صُدُور النَّاس
" [ النَّاس : 5 ] ."
(القرطبي). وَقَدْ
جَاءَ فِي
رُؤْيَتِهِمْ
أَخْبَارٌ صَحِيحَةٌ
(إِذَا
نُقِلُوا
عَنْ
صُوَرِهِمْ)،
ومنها ما رواه
أبو هريرة: (وكَّلني رسولُ
اللهِ بحفظ
زكاةِ رمضانَ
، فأتانى آتٍ ،
فجعل يحثو من
الطعامِ ،
فأخذتُه ،
فقلتُ :
لأَرفعنَّك
إلى رسولِ اللهِ
، قال : إني
محتاجٌ ،
وعليَّ دَينٌ
وعِيالٌ ، ولي
حاجةٌ شديدةٌ
فخلَّيتُ عنه
، فأصبحتُ،
فقال
النَّبيُّ: يا
أبا هريرةَ ما
فعل أسيرُك البارحةَ؟
قال : قلتُ : يا
رسولَ اللهِ
شكا حاجةً
شديدةً
وعِيالًا ،
فرحمتُه
فخلَّيتُ
سبيلَه ، قال :
أما إنه قد
كذبَك وسيعود
فعرفت أنه سيعودُ
، لقولِ رسولِ
اللهِ : أنه
سيعود ، فرصدتُه
، فجاء يحثو
من الطعامِ (
وذكر الحديثَ
إلى أن قال : )
فأخذتُه (
يعني في
الثالثةِ )
فقلتُ :
لأَرفعنَّكَ
إلى رسولِ
اللهِ ، و هذا آخرُ
ثلاثِ مراتٍ تزعم
أنك لا تعود ،
ثم تعود ، قال :
دَعْني أُعلِّمْك
كلماتٍ ينفعك
اللهُ بها
قلتُ : ما هنَّ
؟ قال ، إذا
أَوَيتَ إلى
فراشِك ،
فاقرأ آيةَ
الكرسيِّ : ( اللهُ
لَا إِلَهَ
إِلَّا هُوَ
الْحَيُّ الْقَيُّومُ
) حتى تختم
الآيةَ ، فإنك
لن يزال عليك
من الله حافظٌ
، ولا يقربُك
شيطانٌ حتى
تصبحَ
فخلَّيتُ
سبيلَه ، فأصبحتُ
، فقال لي رسولُ
اللهِ : ما فعل أسيرُك
البارحةَ ؟
قلتُ : يا
رسولَ اللهِ
زعم أنه
يُعلِّمُني كلماتٍ
ينفعني اللهُ
بها ،
فخلَّيتُ
سبيلَه ، قال:
ما هي؟ قلتُ :
قال لي : إذا
أوَيتَ إلى
فراشِك
فاقرأْ آيةَ
الكُرسيِّ ، من
أولها حتى
تختم الآيةَ (
اللهُ لَا
إِلَهَ إِلَّا
هُوَ الْحَيُّ
الْقَيُّومُ )
، و قال لي : لن
يزال عليك من
الله حافظٌ ،
و لا يقربُك
شيطانٌ حتى
تصبحَ و كانوا
أحرصَ شيءٍ
على الخير
فقال النبيُّ
: أما إنه قد
صدَقَك ، و هو
كذوبٌ ، تعلم
مَن تخاطبُ
منذ ثلاثِ ليالٍ
يا أبا هريرةَ
؟ قلتُ : لا قال :
ذاك الشيطانُ) (الراوي: أبو هريرة المحدث:الألباني - المصدر: صحيح
الترغيب -
الصفحة أو
الرقم:
610، خلاصة
حكم المحدث: صحيح).
الفرع
الثالث: ولو
قرأتَ لأصبحتْ
يراها الناسُ
ما تستتِرُ
منهم
- (أنَّ
أسيدَ بنَ
حضيرٍ ، بينما
هو ، ليلةً ،
يقرأُ في
مربدِه . إذ جالتْ
فرسُه . فقرأ
. ثم جالتْ
أخرى . فقرأ . ثم
جالتْ أيضًا .
قال أسيدٌ :
فخشيتُ أن
تطأَ يحيى .
فقمتُ إليها .
فإذا مثلُ
الظُلَّةِ
فوقَ رأسي .
فيها أمثالُ السُّرُجِ
. عرجت في
الجوِّ حتى ما
أراها . قال فغدوتُ
على رسولِ
اللهِ صلَّى اللهُ
عليهِ
وسلَّمَ
فقلتُ : يا
رسولَ اللهِ ! بينما
أنا البارحةُ
من جوفِ الليلِ
أقرأُ في
مِربدي . إذ
جالت فرسي .
فقال رسولُ
اللهِ صلَّى
اللهُ عليهِ
وسلَّمَ "
اقرأ . ابنَ
حضيرٍ ! " قال :
فقرأتُ . ثم
جالت أيضًا .
فقال رسولُ
اللهِ صلَّى
اللهُ عليهِ
وسلَّمَ " اقرأ
. ابنَ حضيرٍ ! "
قال : فقرأتُ .
ثم جالت أيضًا
. فقال رسولُ
اللهِ صلَّى اللهُ
عليهِ
وسلَّمَ "
اقرأ . ابنَ
حضيرٍ ! " قال
فانصرفتُ .
وكان يحيى
قريبًا منها .
خشيتُ أن تطأَه
. فرأيتُ مثلَ
الظُلَّةِ .
فيها أمثالُ
السُّرُجِ .
عرجتْ في
الجوِّ حتى ما
أراها . فقال
رسولُ اللهِ
صلَّى اللهُ
عليهِ
وسلَّمَ " تلك
الملائكةُ
كانت تستمعُ
لك . ولو قرأتَ لأصبحتْ
يراها الناسُ
. ما تستتِرُ
منهم
" .) (
الراوي: أبو
سعيد الخدري
المحدث:مسلم
-
المصدر: صحيح مسلم -
الصفحة أو
الرقم:
796،
خلاصة حكم
المحدث: صحيح).
يتضح
من هذا الحديث
الشريف أنّ
رؤية أسيدَ
بنَ حضيرٍ
للملائكة
كانت كرامة له
لما قرأه من
القرآن في جوف
تلك اللّيلة. فإنّ
رؤيته هذه
للملائكة
كانت استثناءً
خصّه اللهُ به
بدليل : (ولو
قرأتَ
لأصبحتْ يراها
الناسُ . ما
تستتِرُ منهم ). فالأصل
أنّ الملائكة
تستتِرُ
من الناس ولا
نستطيع رؤيتها
على هيئتها .
- (وحدثنا
يحيى
بن يحيى أخبرنا
أبو
خيثمة عن أبي
إسحق عن البراء قال
(كان رجل
يقرأ سورة
الكهف وعنده
فرس مربوط بشطنين
فتغشته سحابة
فجعلت تدور
وتدنو وجعل
فرسه ينفر
منها فلما
أصبح أتى
النبي صلى
الله عليه
وسلم فذكر ذلك
له فقال تلك
السكينة
تنزلت للقرآن)
(البخاري: باب
نزول السكينة
لقراءة
القرآن ، ص 548 ،
رقم 795).
- (Meanwhile a companion of the Prophet reading Al-Kahf surah and beside him a horse hitched
with two ropes, a rotating cloud starts
approaching him and make his horse alienate.
When the day light appears, he went to the Prophet - peace be upon him -
and mentioned that to him. Prophet
said those tranquility descend for the Qur-an. According to another version, these angels
were listening to you. Had you
read more, it will become apparent to people, and no more hidden).
الفرع
الرابع: هنالك
ما لا يبصرهُ
الإنسان
(فَلَا
أُقْسِمُ
بِمَا
تُبْصِرُونَ *
وَمَا لَا
تُبْصِرُونَ * إِنَّهُ
لَقَوْلُ
رَسُولٍ
كَرِيمٍ) (الحاقة
س 69 : 38-40)
(So I do call to witness what ye see *
And what ye see not * That this is verily the word of an honoured Messenger) (S. 69, V.
38-40)
أورد
ابن كثير في
تفسيره :
(يَقُول
تَعَالَى
مُقْسِمًا
لِخَلْقِهِ
بِمَا
يُشَاهِدُونَهُ
مِنْ آيَاته
فِي مَخْلُوقَاته
الدَّالَّة
عَلَى
كَمَالِهِ
فِي أَسْمَائِهِ
وَصِفَاته.
وَمَا غَابَ
عَنْهُمْ
مِمَّا لَا
يُشَاهِدُونَهُ
مِنْ الْمُغَيَّبَات
عَنْهُمْ
إِنَّ
الْقُرْآن كَلَامه
وَوَحْيه وَتَنْزِيله
عَلَى عَبْده
وَرَسُوله
الَّذِي
اِصْطَفَاهُ
لِتَبْلِيغِ
الرِّسَالَة
وَأَدَاء
الْأَمَانَة
فَقَالَ
تَعَالَى " فَلَا
أُقْسِم
بِمَا
تُبْصِرُونَ
وَمَا لَا تُبْصِرُونَ"
. (إِنَّهُ
لَقَوْلُ
رَسُولٍ
كَرِيمٍ( يَعْنِي
مُحَمَّدًا
صَلَّى
اللَّه
عَلَيْهِ
وَسَلَّمَ
أَضَافَهُ
إِلَيْهِ
عَلَى
مَعْنَى التَّبْلِيغ
لِأَنَّ
الرَّسُول
مِنْ شَأْنه
أَنْ
يُبَلِّغ
عَنْ
الْمُرْسَل
وَلِهَذَا
أَضَافَهُ
فِي سُورَة
التَّكْوِير
إِلَى الرَّسُول
الْمَلَكِيّ
" إِنَّهُ
لَقَوْل
رَسُول
كَرِيم ذِي
قُوَّة عِنْد
ذِي الْعَرْش
مَكِين مُطَاع
ثَمَّ
أَمِين"
وَهَذَا
جِبْرِيل
عَلَيْهِ
السَّلَام
ثُمَّ قَالَ
تَعَالَى "
وَمَا صَاحِبكُمْ
بِمَجْنُونٍ
" يَعْنِي
مُحَمَّدًا
صَلَّى
اللَّه
عَلَيْهِ
وَسَلَّمَ "
وَلَقَدْ
رَآهُ
بِالْأُفُقِ
الْمُبِين "
(التكوير آية 23) يَعْنِي
أَنَّ
مُحَمَّدًا
رَأَى
جِبْرِيل عَلَى
صُورَته
الَّتِي
خَلَقَهُ
اللَّه عَلَيْهَا
" وَمَا هُوَ
عَلَى
الْغَيْب
بِضَنِينٍ "
أَيْ
بِمُتَّهَمٍ
" وَمَا هُوَ
بِقَوْلِ
شَيْطَان
رَجِيم " .
وفي
الحديث
الشريف :
(يا
عائشةُ هذا
جبريلُ يقرأُ
عليك السلامَ
. قالت : قلتُ : وعليه
السلامُ
ورحمةُ اللهِ
، ترى ما لا
نرى ، تريدُ
رسولَ اللهِ
صلى اللهُ عليهِ
وسلَّمَ .) (الراوي : عائشة أم
المؤمنين ، المحدث : البخاري ، المصدر : صحيح
البخاري ، الصفحة
أو الرقم: 6249 ،
خلاصة
حكم المحدث : [أورده
في صحيحه]
وقال : تابعه
شعيب. وقال
يونس
والنعمان عن
الزهري: (وبركاته) ، انظر شرح
الحديث رقم 4731 (
(أن
النبيَّ صلى
الله عليه
وسلم قال لها:
يا عائشةُ ،
هذا جبريلُ
يقرأُ عليكِ
السلامَ. فقالت:
وعليه
السلامُ
ورحمةُ اللهِ
وبركاتُه، ترى
ما لا أرى،
تريدُ
النبيَّ صلى
الله عليه وسلم.) (الراوي : عائشة أم
المؤمنين ، المحدث : البخاري ، المصدر : صحيح
البخاري ، الصفحة
أو الرقم: 3217،
خلاصة
حكم المحدث : [صحيح[ ، شرح
الحديث :
قال
النَّبيُّ
صلَّى الله
عليه وسلَّم:
يا عائشةُ،
هذا جبريلُ
يَقرَأُ
عليكِ
السَّلامَ،
أي: يُهديكِ
السَّلامَ،
ويُحيِّيكِ
بتحيَّةِ
الإسلامِ،
فقالتْ: وعليه
السَّلامُ
ورحمةُ الله
وبركاتُه، أي:
ردَّتِ
التَّحيَّةَ
بأحسَنَ
منها، ثمَّ
قالتْ: تَرى
ما لا أَرى؛
تُريدُ
النَّبيَّ
صلَّى الله عليه
وسلَّم، أي:
إنَّك يا
رسولَ الله،
ترى جبريلَ
الذي لا أَراه.
وفي
الحديثِ:
فضيلةٌ
ظاهرةٌ لأمِّ
المؤمنين
عائشةَ رضي
الله عنها.
وفيه:
بَعْثُ
السَّلام
وتبليغُه،
وبَعْثُ الأجنبيِّ
السَّلامَ
إلى
الأجنبيَّةِ
الصَّالحةِ
إذا لم يُخُفْ
ترتُّبُ
مَفْسَدةٍ.
(يا عائشُ !
هذا جبريلُ
يقرأُ عليك
السلامَ . قالت
فقلت : وعليه
السلامُ
ورحمةُ اللهِ
. قالت : وهو يرى
ما لا أرى .) (الراوي : عائشة أم
المؤمنين ، المحدث : مسلم ، المصدر : صحيح مسلم ، الصفحة
أو الرقم: 2447 ،
خلاصة
حكم المحدث : صحيح ، انظر شرح
الحديث رقم 4731 (
(إنَّ
جبريلَ يقرأُ
عليك
السَّلامَ ،
قالت : وعليه
السَّلامُ
ورحمةُ اللهِ
وبركاتِه ،
ترَى ما لا
نرَى) (الراوي : عائشة أم
المؤمنين ، المحدث : الألباني ، المصدر : صحيح
النسائي ، الصفحة
أو الرقم: 3963 ،
خلاصة
حكم المحدث : صحيح ، انظر شرح
الحديث رقم 4731 (
Many species can see frequencies outside the visible spectrum:
العديد
من أنواع
الحيوانات يمكنه
أن يرى ويبصر
تردّدات خارج الطيف
المرئي:
النحل
وكذلك العديد
من الحشرات
ترى الضوء فوق
البنفسجي. وفي بحث
النحلة عن
الرحيق ؛ فإنّها
ترى الزهور في
الطيف فوق
البنفسجي أجمل
من رؤية
الإنسان
للزهور في
الطيف المرئي. وكذلك
الطيور
فإنّها ترى في
الطيف فوق
البنفسجي (300–400 nm)
، ومع
تزايد الطول
الموجي ؛ فإنّ
قدرة الطيور على
الإبصار
تتوقّف عند
الطول الموجي
(590nm) ؛
أي قبل بداية
اللّون
البرتقالي
مباشرة. وترى
الطيور نفسها
أكثر جمالاً
ممّا تبدو
للإنسان ،
وذلك لوجود
علامات على
ريشها لا تظهر
إلا في الطيف
فوق البنفسجي.
Many species can see light with frequencies outside the
"visible spectrum," which is defined in terms of human vision. Bees and many other insects can detect ultraviolet light, which
helps them find nectar in flowers. Plant species that depend on insect pollination
may owe reproductive success to their appearance in ultraviolet light, rather
than how colorful they appear to humans. Birds, too, can see into the
ultraviolet (300–400 nm), and some have sex-dependent markings on their
plumage (ريشها) that are visible only in the ultraviolet range.[2][3] Many animals that can see into the ultraviolet range,
however, cannot see red light or any other reddish wavelengths. Bees' visible spectrum ends at about
590nm, just before the orange wavelengths start. Birds, however, can see some red
wavelengths, but not as many as humans. The common goldfish is the only animal
that can see both infrared and ultraviolet light. (http://en.wikipedia.org/wiki/Visible_spectrum).
المبحث الثاني : الرؤية الليلية
Night
vision
(From
Wikipedia, the free encyclopedia :
http://en.wikipedia.org/wiki/Night_vision)
Night
vision is the ability to see in low light conditions. Whether by
biological or technological means, night vision is made possible by a
combination of two approaches: sufficient spectral range, and sufficient
intensity range. Humans have poor night vision compared to many animals, in
part because the human eye lacks a tapetum lucidum.[1]
تتطلّب
الرؤية
الليلية
كفاية في كلّ
من : سعة المدى
الطيفي ،
وكذلك كفاية
في شدّة
الطيف.
Types of ranges (أنواع
من النطاقات)
Spectral range (المدى
الطيفي) :
Night-useful
spectral range techniques can sense radiation that is invisible to a human
observer. Human vision is confined to a small portion of the electromagnetic spectrum called visible light. Enhanced spectral range allows the
viewer to take advantage of non-visible sources of electromagnetic radiation
(such as near-infrared or ultraviolet radiation). Some animals can see using much more of the infrared and/or
ultraviolet spectrum than humans.
بعض
الحيوانات
يمكنها
الرؤية على
نطاق واسع من
الأطوال
الموجيّة (من
الأشعة تحت
الحمراء إلى
الأشعة فوق
البنفسجيّة) .
وبالتالي
فإنّها تستقبل
وتوظّف كمية كافية
من شدّة الطيف ؛
فترى في
الظلام
الحالك.
Intensity range (مدى
شدّة الإشعاع)
Sufficient
intensity range is simply the ability to see with very small quantities of
light.[2]
Many
animals have better night vision than humans do, the result of one or more
differences in the morphology and anatomy of their eyes. These include having a
larger eyeball, a larger lens, a larger optical aperture (the pupils may expand to the physical limit of the eyelids),
more rods than cones (or rods exclusively) in the retina, and a tapetum lucidum.
Enhanced
intensity range is achieved via technological means through the use of an image intensifier, gain multiplication CCD,
or other very low-noise and high-sensitivity array of photodetectors.
Biological night vision (الرؤية
الليلية الحيوية)
In
biological night vision, molecules of rhodopsin in the rods of the eye undergo a change in shape as they absorb light. Rhodopsin is
the chemical that allows night-vision, and is extremely sensitive to light.
Exposed to a spectrum of light, the pigment immediately bleaches, and it takes
about 30 minutes to regenerate fully, but most of the adaptation occurs within the first five or ten minutes in the dark.
Rhodopsin in the human rods is less sensitive to the longer red wavelengths of light, so traditionally many people use red light to help
preserve night vision as it only slowly depletes the eye's rhodopsin stores in
the rods and instead is viewed by the cones. However the US submarine force ceased using red lighting for night adaptation after
studies found little significant advantage of using low level red over low
level white lighting.[3] [4] Many animals have a tissue layer called the tapetum lucidum
in the back of the eye that reflects light back through the retina, increasing the
amount of light available for it to capture. This is found in many nocturnal animals (activity during the
night and sleeping during the day) and some deep sea animals, and is the cause of eyeshine. Humans lack a tapetum
lucidum.
كثير
من الحيوانات
الليلية
لديها بساط من
طبقة أنسجة (tapetum lucidum) تقع
في الجزء
الخلفي من
العين (بينما
لا توجد هذه
الطبقة لدى
الإنسان). يقوم
هذا البساط بعكس
الضوء إلى
شبكية العين،
مما يزيد من
كمية الضوء المتاحة
للالتقاط. كما وتوجد
طبقة الأنسجة
هذه في أعين بعض
الحيوانات التي
تعيش في أعماق
البحار. تسهم
هذه الطبقة في
زيادة شدة
الضوء المتاح
في النظام البصري
، ممّا يسمح
بالرؤية الليلية
رغم الإضاءة
المنخفضة . وهذا
يفسّر السبب في
لمعان عيون هذه
الحيوانات في
اللّيل.
Nocturnal
mammals have rods with unique properties that make enhanced night vision
possible. The nuclear pattern of their rods changes shortly after birth to
become inverted. In contrast to contemporary rods, inverted rods have heterochromatin in the center of their nuclei and euchromatin and other transcription factors along the border. In
addition, the outer nuclear layer (ONL) in nocturnal
mammals is thick due to the millions of rods present to process the lower light
intensities of a few photons. Rather than being
scattered, the light is passed to each nucleus individually.[5] In fact, an animal's ability to see in low light levels may
be similar to what humans see when using first- or perhaps second-generation image intensifiers.
---------------------------------
Infrared detection in
animals (رؤية
الأشعة تحت
الحمراء لدى
الحيوانات)
(http://www.mapoflife.org/topics/topic_311_Infrared-detection-in-animals/)
Categories: Arthropods: Insects, Mammals, Reptiles, Senses
التصنيفات:
المفصليات،
الحشرات ، والثدييات
، والزواحف، و
حواسها
Some snakes are famous for 'seeing'
infrared, but did you know that their heat-sensing abilities are rivalled by (ينافسها) some beetles (الخنافس) that can detect forest fires over
considerable distances?
Vision
is defined by its access to the visible electromagnetic spectrum. Many animals,
notably insects and birds, can see into the ultraviolet, although this comes with
the joined dangers that these short wavelengths (10-400 nm) are high energy and
potentially biologically destructive. At the opposite end of the visible
spectrum are orange and red, and there has been considerable speculation
whether any optical system could register infrared, drawing upon a suitably
adapted opsin. The reason to be sceptical
(يكون
متشككا) is
that the radiation of these long wavelengths (750 nm-1 mm) is of low energy,
and indeed to date there is no direct evidence for infrared vision. We can, of course, sense
it as radiant heat, but several times animals have independently evolved
systems of infrared detection that in a number of respects are closely
analogous to the eye. In a variety of
organisms, transducers (محولات
الطاقة)
process incoming infrared radiation into a sensory signal that is interpreted
by the brain and in at least one case (the snakes) is integrated with visual
information. Obviously, infrared detection is ultimately involved with heat
sources, but these can vary from 'warm-blooded' prey (snakes, vampire bats (الخفافيش
مصّاصة الدماء) and bed bugs) to forest fires
(three independent examples in the beetles (الخنافس)).
Infrared detection in
vertebrates (رؤية
الأشعة تحت
الحمراء لدى
الفقاريات)
Snakes (الأفاعي)
Infrared
detection is probably best known from the snakes, where thermosensitive pits on
the head have evolved at least twice  - once in the pit vipers (Crotalinae) and probably once in the more ancient boas and pythons (which in some taxonomies are united in the family Boidae).
The infrared-sensing pit organs of boids and crotalines are similar with regard
to their ultra structure and electrophysiological function, but differ in
number, location and overall morphology. While the three or more labial pits of
boids are relatively simple, the single loreal pair of crotalines that is located
between eye and nostril has a more complex structure. It follows the
design of a bolometer, with a thin membrane suspended above a lower air-filled
chamber. In both pit types, the heat-sensitive membrane is highly vascular and
innervated with sensory dendrites, which are formed from terminal masses of the
trigeminal nerve. A recent study has suggested that these nerve fibres are
richly endowed with presumed infrared sensors, heat-sensitive ion channels of
the TRPA1 type (orthologues of which mediate chemosensation in mammals and
other vertebrates). Thus, infrared detection is probably indirect, involving a
thermo transduction mechanism, rather than a direct photochemical reaction
analogous to the activation of opsins in the vertebrate eye. Still snakes
basically 'see' infrared, as the thermal signals are combined with visual
information in the optic tectum. The localisation of 'warm-blooded' prey was
long assumed to be the sole function of infrared detection in snakes, but heat
pits seem to be more general-purpose organs that might also be used for
behavioural thermoregulation and perhaps predator detection or den site
selection.
- once in the pit vipers (Crotalinae) and probably once in the more ancient boas and pythons (which in some taxonomies are united in the family Boidae).
The infrared-sensing pit organs of boids and crotalines are similar with regard
to their ultra structure and electrophysiological function, but differ in
number, location and overall morphology. While the three or more labial pits of
boids are relatively simple, the single loreal pair of crotalines that is located
between eye and nostril has a more complex structure. It follows the
design of a bolometer, with a thin membrane suspended above a lower air-filled
chamber. In both pit types, the heat-sensitive membrane is highly vascular and
innervated with sensory dendrites, which are formed from terminal masses of the
trigeminal nerve. A recent study has suggested that these nerve fibres are
richly endowed with presumed infrared sensors, heat-sensitive ion channels of
the TRPA1 type (orthologues of which mediate chemosensation in mammals and
other vertebrates). Thus, infrared detection is probably indirect, involving a
thermo transduction mechanism, rather than a direct photochemical reaction
analogous to the activation of opsins in the vertebrate eye. Still snakes
basically 'see' infrared, as the thermal signals are combined with visual
information in the optic tectum. The localisation of 'warm-blooded' prey was
long assumed to be the sole function of infrared detection in snakes, but heat
pits seem to be more general-purpose organs that might also be used for
behavioural thermoregulation and perhaps predator detection or den site
selection.
Vampire bats
(الخفافيش مصّاصة
الدماء)
 There is some evidence
for an independently evolved capacity to detect infrared radiation in vampire bats . These mammals cut up the skin of other vertebrates with
their razor-sharp teeth to lick up the blood. They evidently locate their
'warm-blooded' prey with the help of three heat-sensitive pits on their nose
that are thermally insulated from the surrounding tissue. A specific nucleus in
the brain seems to have important similarities in terms of histology and location
to the equivalent infrared nucleus found in snakes.
There is some evidence
for an independently evolved capacity to detect infrared radiation in vampire bats . These mammals cut up the skin of other vertebrates with
their razor-sharp teeth to lick up the blood. They evidently locate their
'warm-blooded' prey with the help of three heat-sensitive pits on their nose
that are thermally insulated from the surrounding tissue. A specific nucleus in
the brain seems to have important similarities in terms of histology and location
to the equivalent infrared nucleus found in snakes.
Infrared detection in
insects (رؤية
الأشعة تحت
الحمراء لدى
الحشرات)
In
terms of convergence, the insects provide a far more striking example than the vertebrates,
because the capacity to detect infrared has evolved several times in this
group. Analogous to snakes and vampire bats, the blood-sucking bed bugs (e.g. in the genera Cimex and Leptocimex)
employ this sensory modality to help locate their 'prey', evidently using a
cave-like organ situated on the antennae. Amongst the
hymenopterans, a parasitoid braconid wasp possesses a peculiar type of antennal
sensillum that is inferred to be a wave-guide for infrared perception and might
play a role in finding a potential host.
Amongst the
hymenopterans, a parasitoid braconid wasp possesses a peculiar type of antennal
sensillum that is inferred to be a wave-guide for infrared perception and might
play a role in finding a potential host.
==================
Night
Vision in Animals (الرؤية
الليلية لدى
الحيوانات)
(http://dmohankumar.wordpress.com/2010/04/12/night-vision-in-animals/)
The Night time
world is never closed for nocturnal animals. Unlike humans, many animals
possess adaptations that allow them to see even when the night
sky is very dark. Animals have developed amazing adaptations to
their environments. Most nocturnal animals have large eyes relative to their
body size. This adaptation, along with the ability to dilate their pupils far
wider than humans can, maximizes the amount of light entering into the eye.

Night vision is the
ability to see in a dark environment. Night vision is mainly due to two
adaptive features: Sufficient spectral
range and Sufficient intensity range.
Spectral range
Human vision is confined
to a small portion of the electromagnetic spectrum called visible spectrum. It extends between 450 and 750
nanometers wavelengths. Below 450 nm is the Ultraviolet spectrum and
above 750 nm is the Infrared
spectrum. Some
animals can see using the infrared or ultraviolet spectrum.
Intensity range
Sufficient intensity
range is the ability to sense very small quantities of light. Although
humans can detect single photons under ideal conditions,
the neurological noise filters limit sensitivity to a few tens of
photons. The night vision of animals is due to one or more differences in the
morphology and anatomy of their eyes. These adaptations include large
eyeball, large lens, large optical aperture, more rods in
the retina, presence of a tapetum lucidum, etc.
Large eyes in Nocturnal
Animals
Nocturnal animals have
large eyes, wider pupil, large lens and increased retinal surface to
collect more light. Some animal species have evolved tubular eyes as
part of their evolution to collect more light. Many nocturnal
animals cannot move their eyes but they have
extraordinary rotational ability of the neck. For
example. Owls can rotate their neck through 270°. This helps to
increase the night vision.
Some animals have
a spherical lens and widened cornea to compensate for
reduced eye movement. This along with a large cornea increases the
animal’s field of vision. So they can see better in night even without
moving the head.
Physiology of
Night vision
The vertebrate eyes
have Photosensitive cells called Rods and Cones. Rods are elongated cells mainly
confined in the periphery of the retina. These are meant for Dim
vision in low light and for peripheral vision (الرؤية
المحيطية). Rods, are
extremely light sensitive and their sensitivity is about 500
times greater than the sensitivity of cones. Only one
photon is required to stimulate a rod to send a signal to the
brain. Nocturnal mammals have rods with unique properties that
make enhanced night vision possible. The nuclear pattern of
their rods changes shortly after birth to become inverted. Inverted rods
have heterochromatin in the center of their nuclei and euchromatin and
other transcription factors along the border. The
outer nuclear layer in nocturnal mammals is thick due to
the presence of millions of rods present to process the lower light intensities
of a few photons. Light is passed to each nucleus individually.

Cones on the other
had are pointed cells confined in the central part of the
retina. These are meant for Central vision, Bright vision and Colour vision. Rods have photosensitive pigment
called Rhodopsin and cones have Iodopsin.
Role of Rhodopsin
The molecules
of Rhodopsin in the rods undergo a change in shape as light is
absorbed by them. Rhodopsin is the chemical that allows night-vision, and is
extremely sensitive to light. When exposed to light, it
immediately bleaches, and it takes about 30 minutes to regeneratefully.
Most of the adaptation occurs within the first five or ten minutes in the dark.
Rhodopsin is less sensitive to the longer red wavelengths of light.
So many people use red light to preserve night vision.
Role of Tapetum
Many animals have
a tissue layer called the Tapetum lucidum in the back of the eye
that reflects light back through the retina. This increases the
amount of light entering into the retina. This is found in many nocturnal
animals and some deep sea animals. This causes the phenomenon
of eye shine in these animals. Tapetum lucidum is absent in human
eye. The shining of eyes in Dogs and Cats in vehicle head
light is due to this reflective retina.

Vision in Different
groups of Animals
Different types
of visual sense exist in animals to adapt them with the
environment. Here are some examples of how some animals see the outside world.
Dog and Cats - They have night
vision and can see moving objects rather than the stationary ones.
Their eyes are much more sensitive to movement. Dogs and cats are color blind and
see only very pale shades of color but they have better peripheral and night
vision. Tapetum lucidum is also present in them to reflect light into the
retina.
Hawk - Hawk’s
vision is equivalent to 20/5. Normal vision for people is 20/20. Therefore Hawk
can see an object from 20 feet that most people can see from 5 feet.
Snakes – They use their normal eyes
during the daytime to see things. During the day time , a snake’s
vision dependents on the movement of prey. They ignore any prey that is completely
motionless. At night snakes will use Pit Organs to sense infrared
rays. Pit organs can pick up infrared heat signals from warm objects
in their surroundings.
Falcon – Distant vision is high in
Falcon. They can see a 10 cm. object from a distance of 1.5 km.
Bees – Bees can see
light between wavelengths 300 nm & 650 nm and they can see
polarized light.
Ants - Ants
can see polarized light.
Octopus –
Octopus is the
largest Mollusc and its retina contains 20 million photoreceptors.
Their eye has a flicker fusion frequency of 70/sec in bright light. The
pupil of the eye is rectangular in shape.
Fish - Some fish can see
the infrared wavelengths. They have only rods in the
retina. About 25 million rods/sq. mm of the retina. This high density of
photoreceptors helps them to detect the dim Bio luminescence in the ocean
depths.
Fly - The eye
of flies has a flicker fusion rate of 300/sec. Flicker
fusion rate is the frequency with which the “flicker” of an image cannot be
distinguished as an individual events of the electromagnetic spectrum.
Shark – Shark has no retinal
cones, and therefore cannot detect colors. Shark’s eyes are designed to pick up
as much light as possible, in order to see in water . But their vision is not
as acute.
Bats - Bats can
detect warmth of an animal from about 16 cm away using
its “nose-leaf”.
Penguin – Penguin is well adapted to see
under water. They have flat cornea that allows for clear vision
underwater. Penguins can also see into the ultraviolet range.
Not all animals see the
world as humans do. For many animals, the world is seen in fuzzy shades of
gray, or very “washed out” and pale colors. But some animals can see in
total darkness, or even see colors beyond the visual spectrum, that humans have
never seen. Still others can use binocular vision to spot prey from
thousands of feet away.
==================
Publisher Summary
Birds can see ultraviolet (UV) light because, unlike humans,
their lenses and other ocular media transmit UV, and they possess a class of
photoreceptor, which is maximally sensitive to violet or UV light, depending on
the species. Birds have a tetra chromatic color space, as compared to the
trichromacy of humans. Birds, along with some reptiles and fish, also possess
double cones in large numbers and a cone class. This chapter discusses a range
of behavioral experiments, from several species, which show that UV information
is utilized in behavioral decisions, notably in foraging and signaling. Removal
of UV wavelengths affects mate choice even in species that are colorful to
humans. These studies emphasize that avian and human color perceptions are different
and that the use of human color standards, and even artificial lighting, may
produce misleading results. However, genuinely objective measures of color are
available, as are, importantly, models for mapping the measured spectra into an
avian color space
(http://www.sciencedirect.com/science/article/pii/S0065345408601059).
Ultraviolet
vision in birds: What is its function? ([1])
Abstract
Although UV vision was first demonstrated in birds in the early
1970s, its function is still unknown. Here we review the evidence for UV vision
in birds, discuss the special properties of UV light, lay out in detail
hypotheses for the function of UV vision in birds and discuss their
plausibility. The main hypotheses are that UV vision functions: (i) in
orientation, (ii) in foraging and (iii) in signaling. The first receives
support from studies of homing pigeons, but it would be unwise to conclude that
orientation is UV's primary function in all birds. It is especially important
to test the signaling hypothesis because bird plumage often reflects UV and
tests of theories of sexual selection have virtually always assumed that birds
perceive plumage “colours” as humans do. A
priori this assumption is
unlikely to be correct, for unlike humans, birds see in the UV, have at least
four types of cones and have a system of oil droplets which filters light
entering individual cones
(http://www.sciencedirect.com/science/article/pii/004269899490149X).
Ultraviolet
vision and band-colour preferences in female zebra finches, Taeniopygia guttata
·
SARAH HUNT, INNES C. CUTHILL, JOHN P. SWADDLE,
and ANDREW T.D. BENNETT
Abstract
Zebra finches have previously been found to have preferences for
particular colours of both natural and artificial traits among opposite sex con
specifics. For example, in some studies female zebra finches preferred males
wearing red leg bands to orange-banded and unbanded birds and rejected light
green-banded males. In other studies, females also preferred males with red
beaks to orange-beaked males. However, several authors have failed to replicate
these results. We show that females may fail to show a colour preference
because of the absence or removal of ultraviolet light under experimental
conditions. In mate-choice trials, females observing males through filters that
transmitted ultraviolet preferred red-banded males but where females viewed
males through ultraviolet-blocking filters, no such preference was observed.
Further investigation revealed that the lack of a colour preference when
ultraviolet was absent was probably due to the change in overall appearance of
the bird, rather than the change in appearance of the rings themselves. This work
highlights the importance of proper consideration of the sensory capabilities
of animals in experimental design, particularly with regard to the role of
ultraviolet light in avian colour perception (http://www.sciencedirect.com/science/article/pii/S0003347297905406).
Volume 58,
Issue 4, October 1999, Pages 809–815
Preferences for
ultraviolet partners in the blue tit
·
SARAH HUNT, INNES C. CUTHILL, ANDREW T.D. BENNETT, and RICHARD GRIFFITHS
Abstract
The preference of female blue tits, Parus caeruleus, is
correlated with the brightness of the ultraviolet (UV) crest (قمة); there is also assortative mating (زواج
نسقي) with respect to the crest's UV/violet chroma. However, manipulation of plumage reflectance
is necessary to infer a direct causal link between UV plumage (ريش
الطيور) and mate choice. We gave both male and
female blue tits a choice between a UV-reflecting (‘UV+’) partner and a
partner whose UV plumage reflectance had been removed (‘UV−’). Male
blue tits significantly preferred UV+ females. Similarly, female blue
tits tended to prefer UV-reflecting males, but their UV+ preferences were
nonsignificant. Neither sex showed a preference when conspecifics were replaced
by a heterospecific. This study suggests mutual mate choice but male choice may
be more strongly influenced by the visual appearance of potential mates. This
is one of a few studies to show male mate preferences and the first
demonstration of a direct relationship between UV reflectance and male mate
choice (http://www.sciencedirect.com/science/article/pii/S0003347299912149).
Ultraviolet vision and foraging in terrestrial vertebrates
رؤية
الأشعة فوق
البنفسجية
والبحث عن
الطعام في
الفقاريات
الأرضية
Johanna Honkavaara, Minna
Koivula, Erkki Korpimäki, Heli
Siitari, and Jussi Viitala
Tetrachromatic colour vision, based on four
‘main’ colours and their combinations, is probably the original colour vision
in terrestrial vertebrates. In addition to human visible waveband of light (400–700
nm) and three main colours, it also includes the near ultraviolet part of light
spectrum (320–400 nm). The ecological importance of ultraviolet (UV) vision in
animals has mainly been studied in the context of intra- and inter-sexual signaling,
but recently the importance of UV vision in foraging has received more
attention.
Foraging animals may use either UV cues
(reflectance or absorbance) of food items or UV cues of the environment. So
far, all diurnal birds studied (at least 35 species), some rodents (4 species),
many reptilians (11 species) and amphibians (2 species) are likely able to see
near UV light. This probably allows e.g. diurnal raptors as well as
frugivorous, nectarivorous and insectivorous birds to use foraging cues
invisible to humans. The possible role of UV cues and existing light conditions
should be taken into account when food selection of vertebrate animals is
studied, particularly, in experiments with artificial food items (http://onlinelibrary.wiley.com/doi/10.1034/j.1600-0706.2002.980315.x/abstract?deniedAccessCustomisedMessage=&userIsAuthenticated=false).
Ultraviolet vision, fluorescence and mate choice in a parrot, the budgerigar Melopsittacus undulatus
1.
Sophie M. Pearn*,
2.
Andrew T.D. Bennett and
+Author Affiliations
1.
*Author for
correspondence (sophie.pearn@bristol.ac.uk).
Abstract
As in many parrots, the
plumage of the budgerigar Melopsittacus
undulatus reflects near–ultraviolet (UVA) wavelengths (300–400 nm) and
exhibits UVA–induced fluorescence. However, there have, to our knowledge, been
no tests of whether the yellow fluorescence observed under intense UVA
illumination has any role in signaling. Four experiments were carried out on
wild–type budgerigars, where the presence and absence of UV reflectance and
fluorescence were manipulated using filters. Few studies have attempted to
separate the contribution of UV reflectance to plumage hue as opposed to brightness
or distinguish between a role in sexual as opposed to social preferences.
However, our first experiments show that not only do females consistently
prefer UV–reflecting males, but also that the observed preferences are due to
removal of UV affecting the perceived hue rather than brightness. Furthermore,
we found no effect of the light environment on male response to females,
suggesting that the female preferences relate to plumage colour per se. Whilst UV reflectance
appears important in heterosexual choice by females, it has no detectable
influence on same–sex association preferences. The results from the second
series of experiments suggest that enhancement of the budgerigar's yellow
coloration through fluorescence has no effect on male attractiveness. However,
the fluorescent plumage may play a role in signaling by virtue of the fact that
it absorbs UVA and so increases contrast with nearby UV–reflecting plumage. Our
study provides convincing evidence that UV reflectances can play a role in mate
choice in non–passerines, but no evidence that the yellow fluorescence observed
under UVA illumination is itself important as a signal (http://rspb.royalsocietypublishing.org/content/268/1482/2273.short).
Ultraviolet colour vision and ornamentation in bluethroats
1.
Staffan Andersson and Trond Amundsen
Many birds see in the ultraviolet (300–400 nm),
but there is limited evidence for colour communication (signaling by spectral
shape independently of brightness) in this ‘hidden’ waveband. Such data are
critical for the understanding of extravagant plumage colours, some of which
show considerable UV reflectance. We investigated UV colour vision in female
social responses to the male UV/violet ornament in bluethroats, Luscinia s. svecica. In an
outdoor aviary at the breeding grounds, 16 females were each presented with a
unique pair of males of equal age. In UVR (UV reduction) males, sun-block
chemicals reduced only the UV reflectance and thereby the spectral shape
(colour) of the throat ornament. In NR (neutral reduction) males, an achromatic
pigment in the same base solvent (preen gland fat) was used for a corresponding
but uniform brightness reduction. Both colour and brightness effects were
invisible to human eyes, and were monitored by spectrometry. In 13 of the 16 trials,
the female associated most with the NR male, a preference that implies that UV
colour vision is used in mate choice by female bluethroats. Reflectance
differences between one–year–old and older males were significant only in UV,
suggestive of a UV colour cue in age–related mate preferences.
(http://rspb.royalsocietypublishing.org/content/264/1388/1587.short).
Ultraviolet
reflection and female mate choice in the pied flycatcher, Ficedula hypoleuca
·
Heli Siitari, Johanna Honkavaara, Esa Huhta, Jussi Viitala
Abstract
In pied flycatchers females seem to prefer male territory
quality rather than male characteristics, and the results of female mate choice
experiments are divergent. In this outdoor aviary study, we examined how
altering the ultraviolet reflection of males affects female mate choice
behaviour. We chose pairs of males with similar human-visible dorsal colour and
morphological traits. We then reduced the proportional ultraviolet reflectance
in one male with sunscreen chemicals. The other male was treated with a
chemical that slightly increased the ultraviolet reflectance of the plumage. In
the experiment females clearly preferred males with slightly increased
ultraviolet reflection. Our results indicate that pied flycatcher females use
ultraviolet cues for mate choice when the effect of territory quality is
controlled for. The results give us new information about a possible mechanism
of mate assessment in this species, and indicate the importance of colour cues
in avian mate choice behaviour (http://www.sciencedirect.com/science/article/pii/S0003347201918706).
Part 2:
Some animals
see differently than we do. Some animals, like bees, have cones for colors
we can't see. Some animals have developed a highly-advanced senses of smell
or specialized hearing abilities such as echolocation. Others have acquired
eye adaptations for improved night vision.
ترى
النحلة
ألواناً لا
يراها
الإنسان ، كما
وقد جعل الله
بعض
الحيوانات
مزودة بنظام إبصار
ليلي.
Big
Eyes
The most
interesting feature of nocturnal animals (الحيوانات
التي تخرج
ليلاً) is the size of
their eyes. Large eyes, with a wider pupil, larger lens and increased retinal
surface collect more light. Some animal species have evolved tubular eyes as a
means of increasing their size. Many nocturnal animals cannot move their eyes
within the orbit. Instead, they have evolved extraordinary rotational ability
in the neck. Owls, for example, can rotate their neck through 270° and this
aids their vision.
Some
animals of the night have acquired a spherical lens and widened cornea to
compensate for reduced eye movement. This combined with a wide cornea increases
the animals field of view allowing the head and eyes to remain motionless.
Mirrors
Add Intensity, Eyes glow in the dark
On a dark
night, flash a bright light at your dog or cat's eyes & you notice that
their eyes glow in the dark. It is the tapetum lucidum (meaning "bright
carpet"), an adaptation for night vision. The tapetum is a thick
reflective membrane, 15 cells wide, directly beneath the retina. It collects and re-emits light back to the
retina a second time, giving the rods a second chance to absorb the image
information, thus maximizing the little light available to them. As this light
is reflected off the tapetum, the animal's eyes appear to glow.
Although
nocturnal animals see mostly crude or imperfect shapes, outlines and no colors,
by maximizing their sensitivity to low light levels with the above adaptations,
it is enough for them to hunt, feed and survive in the dark of night.
In The
Daylight
Most
nocturnal animals are often inactive during the day to avoid over-stimulating
their highly sensitive eyes. Nocturnal animals have specialized pupils to shut
out damaging bright day light. Nocturnal animals dilate their pupils to their
circular maximum at night.
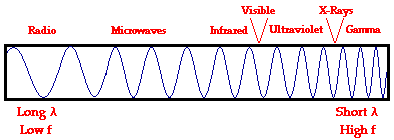
Electromagnetic waves exist with an enormous range of frequencies.
This continuous range of frequencies is known as the electromagnetic spectrum. The entire range of the
spectrum is often broken into specific regions. The subdividing of the entire
spectrum into smaller spectra is done mostly on the basis of how each region of
electromagnetic waves interacts with matter. The diagram below depicts the
electromagnetic spectrum and its various regions. The longer wavelength (![]() ), lower frequency (f) regions are located on the far left of
the spectrum and the shorter wavelength (
), lower frequency (f) regions are located on the far left of
the spectrum and the shorter wavelength (![]() ), higher frequency (f) regions are on the far right. Two very
narrow regions within the spectrum are the visible light region and the X-ray
region.
), higher frequency (f) regions are on the far right. Two very
narrow regions within the spectrum are the visible light region and the X-ray
region.
Visible spectrum (http://en.wikipedia.org/wiki/Visible_spectrum) :
White light dispersed by a prism into the
colors of the optical spectrum.
The visible spectrum
is the portion of the electromagnetic spectrum that is visible to (can be detected by) the human eye.
Electromagnetic radiation in this range of wavelengths is called visible light or
simply light. A typical
human eye will respond to wavelengths from about 390 to 750 nm.[1] In terms of frequency, this
corresponds to a band in the vicinity of 790–400 terahertz (Tera: Is a prefix denoting ![]() ). A light-adapted eye generally has its maximum
sensitivity at around 555 nm (540 THz), in the green region of the optical spectrum (see: luminosity
function). The
spectrum does not, however, contain all the colors that the human eyes and brain can distinguish. Unsaturated
colors such as pink, or purple variations such as magenta, are absent, for example, because
they can only be made by a mix of multiple wavelengths.
). A light-adapted eye generally has its maximum
sensitivity at around 555 nm (540 THz), in the green region of the optical spectrum (see: luminosity
function). The
spectrum does not, however, contain all the colors that the human eyes and brain can distinguish. Unsaturated
colors such as pink, or purple variations such as magenta, are absent, for example, because
they can only be made by a mix of multiple wavelengths.
Visible
wavelengths also pass through the "optical window", the region of the
electromagnetic spectrum that passes largely unattenuated through the Earth's atmosphere. Clean air scatters blue light more than wavelengths
toward the red, which is why the mid-day sky appears blue. The human eye's
response is defined by subjective testing (see CIE),
but atmospheric windows are defined by physical measurement.
The "visible
window" is so called because it overlaps the human visible response
spectrum. The near
infrared (NIR)
windows lie just out of human response window, and the Medium Wavelength IR
(MWIR) and Long Wavelength or Far Infrared (LWIR or FIR) are far beyond the
human response region.
Spectral colors
Colors that can be
produced by visible light of a single wavelength (monochromatic light) are referred
to as the pure
spectral colors.
|
380–450 nm |
668–789 THz |
|
|
450–495 nm |
606–668 THz |
|
|
495–570 nm |
526–606 THz |
|
|
570–590 nm |
508–526 THz |
|
|
590–620 nm |
484–508 THz |
|
|
620–750 nm |
400–484 THz |
 Linear_visible_spectrum.svg (SVG
file, nominally 605 ×
115 pixels, file size: 13 KB)
Linear_visible_spectrum.svg (SVG
file, nominally 605 ×
115 pixels, file size: 13 KB)
(http://upload.wikimedia.org/wikipedia/commons/d/d9/Linear_visible_spectrum.svg)
Although the spectrum is
continuous, with no clear boundaries between one color and the next, the ranges
may be used as an approximation.[9]
Spectroscopy
Rough plot of Earth's atmospheric transmittance (or opacity) to various wavelengths of electromagnetic radiation, including visible light.
Spectroscopy is the study of objects based on the
spectrum of color they emit or absorb. Spectroscopy is an important
investigative tool in astronomy where scientists use it to analyze
the properties of distant objects. Typically, astronomical spectroscopy uses high-dispersion diffraction
gratings to observe
spectra at very high spectral resolutions. Helium was first detected by analyzing the spectrum of the Sun.
Chemical
elements can be detected
in astronomical objects by emission lines and absorption lines. The shifting of spectral lines can
be used to measure the red shift or blue shift of distant or fast-moving objects.
The first exoplanets were discovered by analyzing the Doppler shift of stars at a resolution that
revealed variations in radial velocity as small as a few meters per second. The presence of planets was
revealed by their gravitational influence on the motion of the
stars.
Color display spectrum
Color spectrum generated in a display
device.
Color displays (e.g., computer monitors and televisions) mix red,
green, and blue color to create colors within their respective color triangles, and so can only approximately
represent spectral colors, which are in general outside any color triangle.
.png) No higher resolution available. Spectrum_(brown_background).png (575 × 120 pixels, file size: 3 KB, MIME type:
image/png)
No higher resolution available. Spectrum_(brown_background).png (575 × 120 pixels, file size: 3 KB, MIME type:
image/png)
(http://en.wikipedia.org/wiki/File:Rendered_Spectrum.png)
A render of the color spectrum into
the sRGB color space on a brown background.
Colors outside the color
gamut of the display device result in negative values. If color accurate
reproduction of the spectrum is desired, negative values can be avoided by
rendering the spectra on a gray background. This gives an accurate simulation
of looking at a spectrum on a gray background.[10]
Scanning
The world of desktop
scanners has crossed the threshold of Deep Color where scanners are capable of
capturing a billion or more colors.
Part 3:
Flicker
Sensitivity of the Chicken
Abstract
The photopic
flicker sensitivity of the chicken was determined using an operant conditioning
psychophysical technique. The results show both high- and low-frequency
fall-off in the sensitivity response, which peaked around 15 Hz. Flicker
sensitivity was determined for a range of stimulus luminance levels, and
directly compared to human flicker response measured under similar stimulus
conditions. At five luminance levels (10, 100, 200, 500 and 1000
![]() ), the overall chicken flicker
sensitivity was found to be considerably lower than for humans, except at high
frequencies. A
greater degree of frequency tuning was also found in the chicken response. The critical flicker fusion values were either similar or
slightly higher for chickens compared to humans (40.8, 50.4, 53.3, 58.2 and 57.4 Hz vs 39.2, 54.0, 54.0, 57.4 and 71.5 Hz
respectively for humans and chickens for increasing stimulus luminance level).
A recently proposed model for flicker sensitivity [Vision Research 39 (1999)
533], which incorporates low- and high-pass temporal filters in cascade, was
found to be applicable to the chicken response. From this model, deductions
were made concerning mechanisms controlling the transfer of temporal
information.
), the overall chicken flicker
sensitivity was found to be considerably lower than for humans, except at high
frequencies. A
greater degree of frequency tuning was also found in the chicken response. The critical flicker fusion values were either similar or
slightly higher for chickens compared to humans (40.8, 50.4, 53.3, 58.2 and 57.4 Hz vs 39.2, 54.0, 54.0, 57.4 and 71.5 Hz
respectively for humans and chickens for increasing stimulus luminance level).
A recently proposed model for flicker sensitivity [Vision Research 39 (1999)
533], which incorporates low- and high-pass temporal filters in cascade, was
found to be applicable to the chicken response. From this model, deductions
were made concerning mechanisms controlling the transfer of temporal
information.
Ref.: (http://www.sciencedirect.com/science/article/pii/S0042698901002681)
Vision Res. 2002 Jan;42(1):99-106.
Measuring and modelling the photopic flicker sensitivity of the chicken (Gallus g. domesticus).
Jarvis JR, Taylor NR, Prescott NB, Meeks I, Wathes CM.
Source
Flicker
: اضطرب,
تردد, رمش
measuring and modeling
the photopic flicker sensitivity of the chicken
Figures
See also
Definition
of Contrast: Contrast is the
difference in similarity and comparison to others. It is emphasizing
differences and showing distinctions as in light versus darkness. i.e. contrast sensitivity refers to the ability of the visual
system to distinguish between an object and its background.
|
|
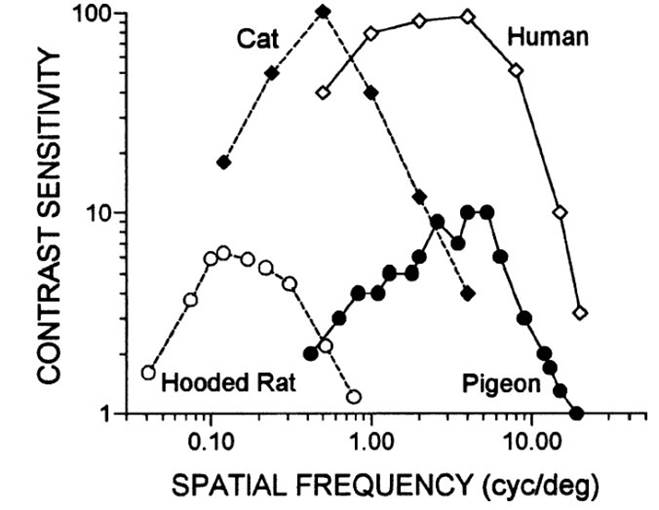
Figure
- The contrast sensitivity function describes how poor the contrast can
become and still be perceptible. As the contrast sensitivity value rises, the
lower the contrast becomes. Combining the experimental data with data for
cats, pigeons, and humans, the hooded rat has comparatively poor eyesight
that may still be useful for medical studies (Ref.: Vision in the Animal
Kingdom: Vision in the Animal Kingdom (Kenneth Kang, Psychology 221 - Applied
Vision and Image Systems, Stanford University - Winter 2002).
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.png){kind=link}
In terms of
the diversity of eyes and vision, birds are quite amazing. While birds have a
structure within their eye called a pecten whose purpose is not clear, they are
capable of seeing ultraviolet and polarized light with about four classes of
cones. Their cone cells have oil droplets to help distinguish colors. Moreover,
they have a higher flicker-fusion frequency, 100 Hz compared to 60 Hz for
humans (Ref.: Vision in the Animal Kingdom).
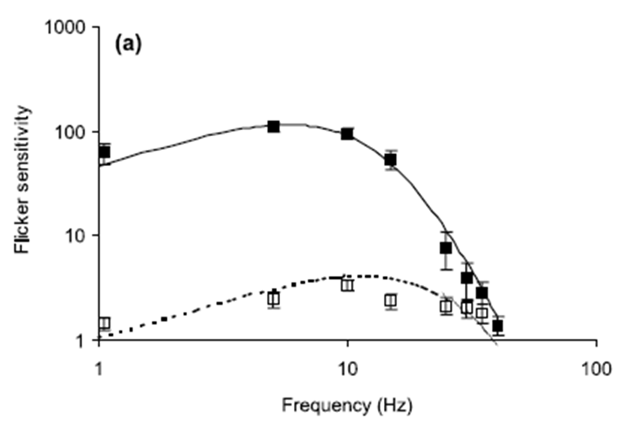
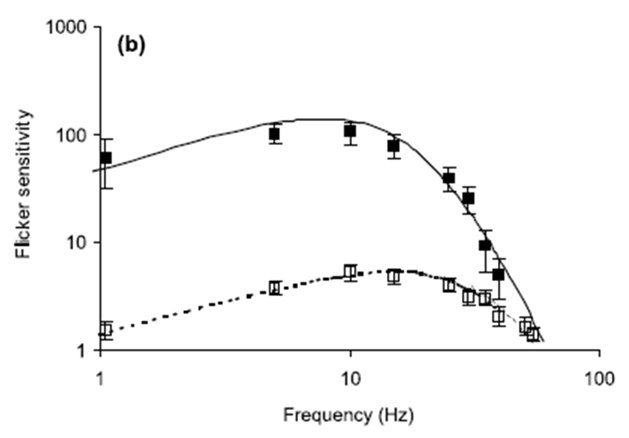
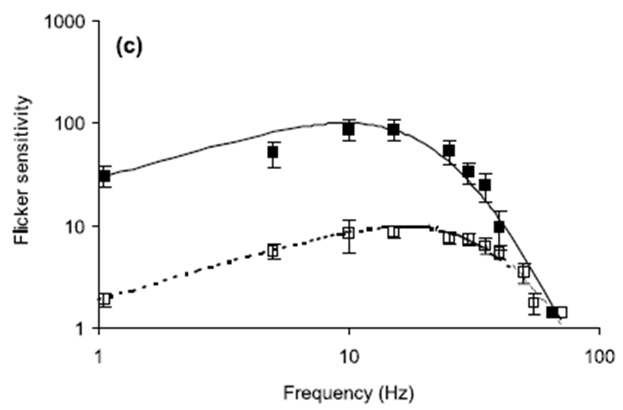
Fig. 1. Flicker sensitivity of chickens and humans. Open
squares show measured chicken data (with SE) while filled squares show measured
human data (with SE). The curves represent flicker sensitivity as determined
from the Rovamo model Eq.
(7). Solid lines are human data and dotted lines are chicken
data. All equation parameters not cited below have values specified in the
text: (a) Stimulus mean luminance; 10 cd/m2. For the human modelling:
N(0)=5.0×10−5 s, Thuman=0.010 s, h=1.3. For the chicken modelling:
N(0)=2.0×10−2 s, Tchicken=0.005 s, h=1.3. (b) Stimulus mean luminance;
200 cd/m2. For the human modelling: N(0)=5.0×10−5 s, Thuman=0.0075 s,
h=1.15. For the chicken modelling: N(0)=3.5×10−2 s, Tchicken=0.0035 s,
h=1.0. (c) Stimulus mean luminance; 1000 cd/m2. For the human modelling:
N(0)=9.0×10−5 s, Thuman=0.006 s, h=1.0. For the chicken modelling:
N(0)=2.0×10−2 s, Tchicken=0.0035 s, h=1.0
Ref.: (http://www.sciencedirect.com/science/article/pii/S0042698901002681)
Ref.: (![]() )
)
References
1.
^ Cecie Starr (2005). Biology: Concepts and Applications. Thomson Brooks/Cole. ISBN 053446226X. http://books.google.com/books?id=RtSpGV_Pl_0C&pg=PA94.
2.
^ Cuthill, Innes C; et al. (1997).
"Ultraviolet vision in birds". in Peter J.B. Slater. Advances in the
Study of Behavior. 29. Oxford, England: Academic Press. p. 161. ISBN 978-0-12-004529-7.
3.
^ Jamieson, Barrie G. M. (2007).
Reproductive Biology and Phylogeny of Birds. Charlottesville VA: University of
Virginia. p. 128. ISBN 1578083869.
4.
^ Coffey, Peter (1912). The Science of Logic: An Inquiry Into
the Principles of Accurate Thought. Longmans. http://books.google.com/books?id=j8BCAAAAIAAJ&pg=PA185&dq=%22roger+bacon%22+prism&ei=TX8OSJ2jMZCSzQTKx8y7Ag&client=firefox-a.
5.
^ Hutchison, Niels (2004). "Music For Measure: On the 300th Anniversary of
Newton's Opticks". Colour Music. http://home.vicnet.net.au/~colmusic/opticks3.htm. Retrieved 2006-08-11.
6.
^ Newton, Isaac (1704). Opticks.
7.
^ Mary Jo Nye (editor) (2003). The Cambridge History of Science: The
Modern Physical and Mathematical Sciences. 5. Cambridge University Press. p. 278. ISBN 9780521571999. http://books.google.com/books?id=B3WvWhJTTX8C&pg=PA278&dq=spectrum+%22thomas+young%22+herschel+ritter&lr=&as_brr=0&as_pt=ALLTYPES&ei=XZT2Se_dF4vOkwT9tMigBA.
8.
^ John C. D. Brand (1995). Lines of light: the sources of
dispersive spectroscopy, 1800-1930. CRC Press. p. 30–32. ISBN 9782884491631. http://books.google.com/books?id=sKx0IBC22p4C&pg=PA30&dq=light+wavelength+color++young+fresnel&as_brr=3&ei=zpX2SdWLIpDmkASaxq3LBA#PPA31,M1.
9.
^ Thomas J. Bruno, Paris D. N.
Svoronos. CRC Handbook of Fundamental
Spectroscopic Correlation Charts. CRC Press, 2005.
10.
^ http://www.repairfaq.org/sam/repspec/
--------------------------
S45.4:
UV vision and its functions in birds
Innes
C. Cuthill, Julian C. Partridge & Andrew T. D. Bennett
School of
Biological Sciences, University of Bristol, Woodland Road, Bristol BS8 1UG, UK,
fax 44 117 925 7374, e-mail I.Cuthill@bris.ac.uk; J.C.Partridge@bris.ac.uk; Andy.Bennett@bris.ac.uk
Cuthill,
I.C., Partridge, J.C. & Bennett, A.T.D. 1999. UV vision and its functions
in birds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol.
Congr., Durban: 2743-2758. Johannesburg:
BirdLife South Africa.
Birds can see
ultraviolet (UV) light because, unlike humans, their lens, cornea and other
ocular media transmit UV, and they possess a retinal cone type which is
maximally sensitive to violet or ultraviolet light, depending on the species.
As birds also have cones sensitive to ‘blue’, ‘green’ and ‘red’ light, they may
have a tetrachromatic colour vision system.
Full article: http://www.int-ornith-union.org/files/proceedings/durban/Symposium/S45/S45.4.htm
--------------------------
Ultraviolet (UV) light perception by
birds: a review, J. Rajchard Faculty of Agriculture, University of South
Bohemia, Ceske Budejovice, Czech Republic.
Veterinarni Medicina, 54, 2009 (8): 351–359: (http://www.vri.cz/docs/vetmed/54-8-351.pdf)
The ability to perceive (observe) the
near ultraviolet part of the light spectrum (the wavelength 320–400 nm) has been detected in many bird species.
It is now known that avian ocular
media do not absorb UV light before it reaches the retina; thus UV sensitivity in birds is possible.
Birds have 4–5 types of single cone photoreceptors, including one type
sensitive to UV light (for comparison humans have only three types of cone
photoreceptors). Many birds (obviously the majority of species, e.g., many non-passerines) have a
violet-sensitive single cone that is obviously sensitive to UV
wavelengths. Other species (e.g., some
passerines) have a single cone that has maximum sensitivity to UV light.
The spectral sensitivity of domestic
ducks (Anas platyrhynchos domesticus) and turkeys (Meleagris gallopavo
gallopavo) was tested over a range of specified wavelengths, including UVA,
between 326–694 nm in comparison with human spectral sensitivity (Barber et
al., 2006). The results showed that ducks and turkeys had similar spectral
sensitivities and could perceive UVA radiation. Turkeys were more sensitive to
UVA than ducks. The peak sensitivity was in the wavelengths between 544–577 nm,
with reduced sensitivity at 508–600 nm. Both bird species had a very different
and broader range of spectral sensitivity than humans.
--------------------------
information. http://hsc.csu.edu.au/biology/options/communication/2950/CommPart2.html
|
Type
of animal |
Name
of animal |
Part
of electromagnetic spectrum detected |
Wavelengths
detected |
|
Vertebrate |
Human |
visible |
700-400
nm |
|
|
Rattlesnake
|
infra-red
and visible |
850-480
nm |
|
|
Japanese
dace fish |
ultraviolet
and visible |
as low as
360 nm |
|
Invertebrate
|
Honeybee |
ultraviolet
and visible |
700-300
nm |
|
|
Mantis
shrimp |
ultraviolet
and visible |
640-400
nm |
Table:
(http://anthonymbiotask3.wikispaces.com/Light+and+the+electromagnetic+spectrum) or
http://hsc.csu.edu.au/biology/options/communication/2950/CommPart2.html
|
Type
of animal |
Name
of animal |
Electromagnetic
spectrum used |
Reasons |
|
Vertebrate
|
Human |
visible |
Active
during the day uses colour for perception of objects |
|
|
Rattlesnake
|
infra-red
and visible |
Active at
night hunts in dark burrows |
|
|
Hummingbird
|
visible |
Can
detect flowers from over a kilometre away |
|
Invertebrate
|
Honeybee |
ultraviolet
and visible |
Can
detect ultraviolet markings on flowers and uses polarised light for
navigation |
|
|
Mantis
shrimp |
ultraviolet
and visible |
Can
perceive many more colours and escape predation in the well lit waters were
it lives |
--------------------------
Part 4: Color vision:
From Wikipedia, the free encyclopedia (http://en.wikipedia.org/wiki/Color_vision)
Color vision is the capacity of an organism or
machine to distinguish objects based on the wavelengths (or frequencies) of the light they reflect, emit, or transmit. The nervous system derives
color by comparing the responses to light from the several types of cone photoreceptors in the eye. These cone
photoreceptors are sensitive to different portions of the visible spectrum. For humans, the visible spectrum
ranges approximately from 380 to 740 nm, and there are normally three types of
cones. The visible range and number of cone types differ between species.
A 'red' apple does not
emit red light.[1] Rather, it simply absorbs all the frequencies of visible light shining on it except for a group of
frequencies that is perceived as red, which are reflected. An apple is
perceived to be red only because the human eye can distinguish between different
wavelengths. The advantage of color, which is a quality constructed by the
visual brain and not a property of objects as such, is the better
discrimination of surfaces allowed by this aspect of visual processing. In some dichromatic substances (e.g. pumpkin seed oil) the color hue
depends not only on the spectral properties of the substance, but also on its
concentration and the depth or thickness[2].
Wavelength and hue detection
Isaac Newton discovered that white light splits
into its component colors when passed through a prism, but that if those bands
of colored light pass through another and rejoin, they make a white beam. The
characteristic colors are, from low to high frequency: red, orange, yellow,
green, cyan, blue, violet. Sufficient differences in frequency give rise to a
difference in perceived hue (hue: is the degree to which a stimulus can be described as similar to
or different from stimuli that are described as red, green, blue, and yellow); the just noticeable difference in wavelength varies from about 1 nm in the blue-green and yellow wavelengths, to 10 nm and more in the red and blue. Though
the eye can distinguish up to a few hundred hues, when those pure spectral
colors are mixed together or diluted with white light, the number of
distinguishable chromaticities can be quite high.
In very low light
levels, vision is scotopic, meaning mediated by rod cells, and not detecting color
differences; the rods are maximally sensitive to wavelengths near 500 nm. In
brighter light, such as daylight, vision is photopic, in which case the cone cells of the retina mediate color perception,
and the rods are essentially saturated; in this region, the eye is most
sensitive to wavelengths near 555 nm. Between these regions is known as mesopic vision, in which case both rods and cones
are providing meaningful signal to the retinal
ganglion cells. The
shift in color perception across these light levels gives rise to differences
known as the Purkinje
effect.
The perception of
"white" is formed by the entire spectrum of visible light, or by
mixing colors of just a few wavelengths, such as red, green, and blue, or even
by mixing just a pair of complementary
colors such as blue
and yellow.[3]
Physiology of color perception

Normalized response
spectra of human cones, short (S), medium (M), and long (L) types, to
monochromatic spectral stimuli, with wavelength given in nanometers.

The same figures as
above represented here as a single curve in three (normalized cone response)
dimensions

Single color sensitivity
diagram of the human eye.
Perception of color is
achieved in mammals through color receptors containing
pigments with different spectral
sensitivities. In
most primates closely related to humans there are three types of color receptors (known as cone cells). This confers trichromatic color vision, so these primates, like humans, are known as trichromats. Many other primates and other
mammals are dichromats, and many mammals have little or no
color vision. Indeed, "mammals with color vision are rare," with most
mammals having rod-dominated retinas, and some having pure-rod ones.[4]
The cones are
conventionally labeled according to the ordering of the wavelengths of the
peaks of their spectral
sensitivities: short
(S), medium (M), and long (L) cone types, also sometimes referred to as blue,
green, and red cones. While the L cones are often referred to as the red
receptors, microspectrophotometry has shown that their peak sensitivity is in
the greenish-yellow region of the spectrum. Similarly, the S- and M-cones do
not directly correspond to blue and green, although they are often depicted as such (such as in the
graph to the right). It is important to note that the RGB color model is merely a convenient means for
representing color, and is not directly based on the types of cones in the
human eye.
The peak response of human
color receptors varies, even amongst individuals with 'normal' color vision;[5] in non-human species this
polymorphic variation is even greater, and it may well be adaptive.[6]
Theories of color vision
Two complementary
theories of color vision are the trichromatic
theory and the opponent process theory. The trichromatic theory, or Young–Helmholtz
theory, proposed in
the 19th century by Thomas
Young and Hermann
von Helmholtz, as
mentioned above, states that the retina's three types of cones are
preferentially sensitive to blue, green, and red. Ewald Hering proposed the opponent process theory
in 1872.[7] It states that the visual system
interprets color in an antagonistic way: red vs. green, blue vs. yellow, black
vs. white. We now know both theories to be correct, describing different stages
in visual physiology.[8]
Cone cells in the human eye
|
Cone
type |
Name |
Range |
|
|
S |
β |
400–500 nm |
420–440 nm |
|
M |
γ |
450–630 nm |
534–545 nm |
|
L |
ρ |
500–700 nm |
564–580 nm |
A range of wavelengths of light stimulates each of these receptor
types to varying degrees. Yellowish-green light, for example, stimulates both L and M cones equally strongly, but
only stimulates S-cones weakly. Red light, on the other hand, stimulates L cones
much more than M cones, and S cones hardly at all; blue-green light stimulates
M cones more than L cones, and S cones a bit more strongly, and is also the
peak stimulant for rod cells; and blue light stimulates almost exclusively S-cones. Violet light
appears to stimulate both L and S cones to some extent, but M cones very
little, producing a sensation that is somewhat similar to magenta. The brain
combines the information from each type of receptor to give rise to different
perceptions of different wavelengths of light.
The pigments present in
the L and M cones are encoded on the X chromosome; defective encoding of these leads
to the two most common forms of color blindness. The OPN1LW gene, which codes for
the pigment that responds to yellowish light, is highly polymorphic (a recent study by Verrelli and
Tishkoff found 85 variants in a sample of 236 men[11]), so up to twenty percent of women[12] have an extra type of color
receptor, and thus a degree of tetrachromatic color vision.[13] Variations in OPN1MW, which codes
for the bluish-green pigment, appear to be rare, and the observed variants have
no effect on spectral
sensitivity.
Color in the human brain

Visual pathways in the
human brain. The ventral
stream (purple) is
important in color recognition. The dorsal stream (green) is also shown. They
originate from a common source in the visual cortex.
Color processing begins
at a very early level in the visual system (even within the retina) through
initial color opponent mechanisms. Opponent mechanisms refer to the opposing
color effect of red-green, blue-yellow, and light-dark. Visual information is
then sent back via the optic nerve to the optic chiasma: a point where the two optic nerves
meet and information from the temporal (contralateral) visual field crosses to
the other side of the brain. After the optic chiasma the visual fiber tracts
are referred to as the optic
tracts, which enter
the thalamus to synapse at the lateral geniculate nucleus (LGN). The LGN is segregated into six layers: two
magnocellular (large cell) achromatic layers (M cells) and four parvocellular
(small cell) chromatic layers (P cells). Within the LGN P-cell layers there are
two chromatic opponent types: red vs. green and blue vs. green/red.
After synapsing at the LGN, the visual tract
continues on back toward the primary visual cortex (V1) located at the back of the
brain within the occipital
lobe. Within V1
there is a distinct band (striation). This is also referred to as "striate
cortex", with other cortical visual regions referred to collectively as
"extrastriate cortex". It is at this stage that color processing
becomes much more complicated.
In V1 the simple
three-color segregation begins to break down. Many cells in V1 respond to some
parts of the spectrum better than others, but this "color tuning" is
often different depending on the adaptation state of the visual system. A given
cell that might respond best to long wavelength light if the light is relatively
bright might then become responsive to all wavelengths if the stimulus is
relatively dim. Because the color tuning of these cells is not stable, some
believe that a different, relatively small, population of neurons in V1 is
responsible for color vision. These specialized "color cells" often
have receptive fields that can compute local cone ratios. Such
"double-opponent" cells were initially described in the goldfish
retina by Nigel Daw;[14][15] their existence in primates was
suggested by David H.
Hubel and Torsten Wiesel and subsequently proven by Bevil
Conway.[16] As Margaret Livingstone and David
Hubel showed, double opponent cells are clustered within localized regions of
V1 called blobs, and are thought to come in two flavors, red-green and
blue-yellow.[17] Red-green cells compare the relative
amounts of red-green in one part of a scene with the amount of red-green in an
adjacent part of the scene, responding best to local color contrast (red next
to green). Modeling studies have shown that double-opponent cells are ideal
candidates for the neural machinery of color constancy explained by Edwin H. Land in his retinex theory.[18]

This image (when viewed
in full size, 1000 pixels wide) contains 1 milion pixels, each of a different
color. The human eye can distinguish about 10 million different colors.[19]
From the V1 blobs, color
information is sent to cells in the second visual area, V2. The cells in V2
that are most strongly color tuned are clustered in the "thin
stripes" that, like the blobs in V1, stain for the enzyme cytochrome
oxidase (separating the thin stripes are interstripes and thick stripes, which
seem to be concerned with other visual information like motion and
high-resolution form). Neurons in V2 then synapse onto cells in the extended
V4. This area includes not only V4, but two other areas in the posterior
inferior temporal cortex, anterior to area V3, the dorsal posterior inferior
temporal cortex, and posterior TEO.[20][21] (Area V4 was identified by Semir Zeki to be exclusively dedicated to
color, but this has since been shown not to be the case.[22] Color processing in the extended V4
occurs in millimeter-sized color modules called globs.[20][21] This is the first part of the brain
in which color is processed in terms of the full range of hues found in color space.[20][21]
Anatomical studies have
shown that neurons in extended V4 provide input to the inferior temporal lobe . "IT" cortex is thought
to integrate color information with shape and form, although it has been
difficult to define the appropriate criteria for this claim. Despite this
murkiness, it has been useful to characterize this pathway (V1 > V2 > V4
> IT) as the ventral
stream or the
"what pathway", distinguished from the dorsal stream ("where pathway") that is
thought to analyze motion, among many other features.
In other animals
Many invertebrates have color vision. Honey- and bumblebees have trichromatic color vision,
which is insensitive to red but sensitive in ultraviolet. Papilio butterflies possess six types of
photoreceptors and may have pentachromatic vision.[23] The most complex color vision system
in animal kingdom has been found in stomatopods (such as the mantis shrimp) with up
to 12 different spectral receptor types thought to work as multiple dichromatic
units.[24].
Vertebrate animals such
as tropical fish and birds sometimes have more complex color vision systems than
humans.[25] In the latter example, tetrachromacy is achieved through up to four cone types, depending on species.
Brightly colored oil droplets inside the cones shift or narrow the spectral
sensitivity of the cell. It has been suggested that it is likely that pigeons are pentachromats.
Reptiles and amphibians
also have four cone types (occasionally five), and probably see at least the
same number of colors that humans do, or perhaps more. In addition, some
nocturnal geckos have the capability of seeing color in dim light[26].
In the evolution of
mammals, segments of color vision were lost, then for a few species of
primates, regained by gene-duplication. Eutherian mammals other than primates (for
example, dogs, cats, mammalian farm animals) generally have less-effective
two-receptor (dichromatic) color perception systems, which
distinguish blue, green, and yellow—but cannot distinguish reds. The adaptation
to see reds is particularly important for primate mammals, since it leads to
identification of fruits, and also newly sprouting leaves, which are
particularly nutritious.
However, even among
primates, full color vision differs between new-world and old-world monkeys.
Old-world primates, including monkeys and all apes, have vision similar to
humans. New
World Monkeys may or
may not have color sensitivity at this level: in most species, males are
dichromats, and about 60% of females are trichromats, but the owl monkeys are cone monochromats, and both sexes of howler monkeys are trichromats.[27][28][29][30] Visual sensitivity differences
between males and females in a single species is due to the gene for
yellow-green sensitive opsin protein (which confers ability to differentiate
red from green) residing on the X sex chromosome.
Several marsupials such as the fat-tailed
dunnart (Sminthopsis
crassicaudata) have been shown to have trichromatic color vision[31].
Marine
mammals, adapted for
low-light vision, have only a single cone type and are thus monochromats.
References
1.
^ Wright, W. D. (1967). The rays are
not coloured: essays on the science and vision and colour. Bristol: Hilger. ISBN 0-85274-068-9.
2.
^ Kreft S and Kreft M (2007)
Physicochemical and physiological basis of dichromatic color,
Naturwissenschaften 94, 935-939. On-line PDF
3.
^ "Eye, human." Encyclopædia
Britannica 2006 Ultimate Reference Suite DVD, 2009.
4.
^ Ali, Mohamed Ather; Klyne, M.A.
(1985). Vision in Vertebrates. New York: Plenum Press. pp. 174-175. ISBN 0-306-42065-1.
5.
^ Neitz J, Jacobs GH (1986). "Polymorphism of the
long-wavelength cone in normal human color vision". Nature 323 (6089): 623–5. doi:10.1038/323623a0. PMID 3773989. http://www.nature.com/nature/journal/v323/n6089/abs/323623a0.html.
6.
^ Jacobs GH (January 1996). "Primate photopigments and
primate color vision". Proc. Natl. Acad. Sci. U.S.A. 93 (2): 577–81. doi:10.1073/pnas.93.2.577. PMID 8570598.
7.
^ Hering, Ewald (1872). "Zur Lehre vom Lichtsinne". Sitzungsberichte der
Mathematisch–Naturwissenschaftliche Classe der Kaiserlichen Akademie der
Wissenschaften LXVI. Band (III Abtheilung). http://books.google.com/books?id=u5MCAAAAYAAJ&pg=PA5&lpg=PA5&dq=1872+hering+ewald+Zur+Lehre+vom+Lichtsinne.+Sitzungsberichte+der+kaiserlichen+Akademie+der+Wissenschaften.+Mathematisch%E2%80%93naturwissenschaftliche+Classe,&source=web&ots=fAdrz1yI8x&sig=99NSKb_P8-_QSDO1RTzt35QTRyk&hl=en.
8.
^ Ali, M.A. & Klyne, M.A. (1985),
p.168
9.
^ Wyszecki, Günther; Stiles, W.S.
(1982). Color Science: Concepts and Methods, Quantitative Data and Formulae
(2nd ed.). New York: Wiley Series in Pure and Applied Optics. ISBN 0-471-02106-7.
10.
^ R. W. G. Hunt (2004). The
Reproduction of Colour (6th ed.). Chichester UK: Wiley–IS&T Series in
Imaging Science and Technology. pp. 11–2. ISBN 0-470-02425-9.
11.
^ Verrelli BC, Tishkoff SA (September
2004). "Signatures of selection and
gene conversion associated with human color vision variation". Am. J. Hum. Genet. 75 (3):
363–75. doi:10.1086/423287. PMID 15252758.
12.
^ [Caulfield HJ (17 April 2006). "Biological color vision inspires artificial
color processing". SPIE Newsroom. doi:10.1117/2.1200603.0099. http://www.spie.org/x8849.xml?highlight=x2410.
13.
^ Roth, Mark (2006). "Some women may see 100 million
colors, thanks to their genes" Post-Gazette.com
14.
^ Nigel W. Daw (17 November 1967).
"Goldfish Retina: Organization for Simultaneous Color Contrast".
Science 158 (3803): 942–4. doi:10.1126/science.158.3803.942. PMID 6054169.
15.
^ Bevil R. Conway (2002). Neural Mechanisms of Color Vision:
Double-Opponent Cells in the Visual Cortex. Springer. ISBN 1402070926. http://books.google.com/books?id=pFodUlHfQmcC&pg=PR7&dq=goldfish+retina+by+Nigel-Daw&as_brr=3&ei=2AWqR764JI7-iAGh8vwE&sig=7vvLHGgrRP_QtPH6mjLuiqblglU.
16.
^ Conway BR (15 April 2001). "Spatial structure of cone
inputs to color cells in alert macaque primary visual cortex (V-1)". J. Neurosci. 21 (8):
2768–83. PMID 11306629. http://www.jneurosci.org/cgi/content/full/21/8/2768.
17.
^ John E. Dowling (2001). Neurons and Networks: An Introduction
to Behavioral Neuroscience. Harvard University Press. ISBN 0674004620. http://books.google.com/books?id=adeUwgfwdKwC&pg=PA376&dq=Margaret+Livingstone+David+Hubel+double+opponent+blobs&as_brr=3&ei=YQaqR9-lAY6CiQHm1cmnCg&sig=D3znxI88shgNd8onK0RAWEMh6zY.
18.
^ McCann, M., ed. 1993. Edwin H. Land's Essays. Springfield, Va.: Society
for Imaging Science and Technology.
19.
^ Judd, Deane B.; Wyszecki, Günter
(1975). Color in Business, Science and Industry. Wiley Series in Pure and
Applied Optics (3rd ed.). New York: Wiley-Interscience. p. 388. ISBN 0471452122.
20.
^ a b c Conway BR, Moeller S, Tsao DY. (2007).
Specialized color modules in macaque extrastriate cortex. Neuron. 56(3):560-73.
PMID 17988638
21.
^ a b c Conway BR, Tsao DY. (2009). Color-tuned
neurons are spatially clustered according to color preference within alert
macaque posterior inferior temporal cortex. Proc Natl Acad Sci U S A.
106:18035-18039. PMID 19805195
22.
^ John Allman and Steven W. Zucker
(1993). "On cytochrome oxidase blobs in
visual cortex".
in Laurence Harris and Michael Jenkin, editors. Spatial Vision in Humans and
Robots: The Proceedings of the 1991 York Conference. Cambridge University
Press. ISBN 0521430712. http://books.google.com/books?id=eWBiKaOCNIYC&pg=PA34&dq=v4+zeki+color&lr=&as_brr=3&ei=KBCqR7eGF4bQiwHpnZSoCg&sig=F_rbsAj3FD69wRMzWGhB1vK4RuQ.
23.
^ Arikawa K (November 2003). "Spectral organization of the
eye of a butterfly, Papilio". J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav.
Physiol. 189 (11): 791–800. doi:10.1007/s00359-003-0454-7. PMID 14520495. http://www.springerlink.com/content/whjepqnhpulyeevk/.
24.
^ Cronin TW, Marshall NJ (1989). "A retina with at least ten
spectral types of photoreceptors in a mantis shrimp". Nature 339: 137–40. doi:10.1038/339137a0. http://www.nature.com/nature/journal/v339/n6220/abs/339137a0.html.
25.
^ Kelber A, Vorobyev M, Osorio D
(February 2003). "Animal color vision—behavioural
tests and physiological concepts". Biol Rev Camb Philos Soc 78 (1): 81–118. doi:10.1017/S1464793102005985. PMID 12620062. http://www.blackwell-synergy.com/doi/abs/10.1017/S1464793102005985.
26.
^ Roth, Lina S. V.; Lundström, Linda;
Kelber, Almut; Kröger, Ronald H. H.; Unsbo, Peter (March 30, 2009). "The pupils and optical systems of gecko eyes". Journal of Vision 9 (3:27):
1–11. doi:10.1167/9.3.27. http://journalofvision.org/9/3/27/.
27.
^ Jacobs, G. H., & Deegan, J. F.
(2001). Photopigments and color vision in New World monkeys from the family
Atelidae. Proceedings of the Royal Society of London, Series B, 268, 695-702.
28.
^ Jacobs, G. H., Deegan, J. F., Neitz,
J., Crognale, M. A., & Neitz, (1993). Photopigments and color vision in the
nocturnal monkey, Aotus. Vision Research, 33, 1773-1783
29.
^ Mollon, J. D., Bowmaker, J. K.,
& Jacobs, G. H. (1984). Variations of color vision in a New World primate
can be explained by polymorphism of retinal photopigments. Proceedings of the
Royal Society of London, Series B, 222, 373-399.
30.
^ Sternberg, Robert J. (2006):
Cognitive Psychology. 4th Ed. Thomson Wadsworth.
31.
^ Arrese CA, Beazley LD, Neumeyer C
(March 2006). "Behavioural evidence for marsupial trichromacy". Curr.
Biol. 16 (6): R193–4. doi:10.1016/j.cub.2006.02.036. PMID 16546067.
--------------------------
The color
temperature of a light source is the temperature of an ideal black body radiator that radiates light of comparable hue
to that of the light source. Color temperature is conventionally stated in the
unit of absolute temperature, the kelvin, having the unit symbol K.
To find the peak of
the blackbody radiation curve, Wien's Displacement Law gives: (http://hyperphysics.phy-astr.gsu.edu/hbase/wien.html)
![]()
If the temperature is
800K, then the wavelength at which the radiation curve peaks is: λpeak
= 6225000000000005 microns
The corresponding
frequency is 82815734989648.03313 Hz.
Planck's constant: h =
6.626068 × 10-34 m2 kg / s.
The corresponding
blackbody radiation has photon energy
hν = 8.28157*6.626068*(10^-21)/(1.6*10^-19)
= 0.342964 eV;
which is in the infrared
range.
|
Name |
Wavelength |
Frequency (Hz) |
Photon Energy (eV) |
|
10 nm -
390 nm |
30 PHZ -
790 THz |
3 to 124 |
|
|
390 nm -
750 nm |
790 THz -
405 THz |
1.7 - 3.3
|
|
|
Infrared |
750 nm - 1 mm |
405 THz - 300 GHz |
0.00124 - 1.7 |
يقول
الله سبحانه
وتعالى: (خَلَقَ
الْإِنْسَانَ
مِنْ
صَلْصَالٍ
كَالْفَخَّارِ
وَخَلَقَ
الْجَانَّ مِنْ
مَارِجٍ مِنْ
نَارٍ). إذا
كانت درجة
حرارة الْجَانَّ
تساوي أو أقل
من 800K ، فان
الأشعة
الصادرة عنه
تكون تحت
الحمراء (hν = 0.342964 eV)
فيتعذر على
الإنسان
رؤيته .
http://how-it-looks.blogspot.com/2010/01/infrared-radiation-black-bodies-and.html
Somewhere in the range
900K to 1000K, the blackbody spectrum encroaches enough in the the visible to be seen as a dull red glow. Most
of the radiated energy is in the infrared. (http://hyperphysics.phy-astr.gsu.edu/hbase/bbrc.html).
![]() To English
To English
![]() To Arabic-English
To Arabic-English
![]() To Arabic
To Arabic